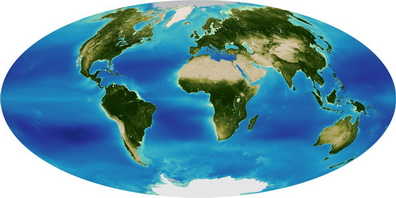
Global
and regional plant distribution
Precipitation, temperature and nutrients control the global distribution
of terrestrial plants, but aquatic plants seem to follow different patterns.
Water chemistry is important and novel analyses reveal exciting patterns of
aquatic plant distribution. More…