
Exogenous ABA can induce a functional root barrier to radial O2 loss in barley
Outer apoplastic barriers in roots – including the barrier to radial O2 loss – can be considered a high-quality Swiss army knife. The reason is that these barriers can restrict the entry of soil phytotoxins (Fe2+, H2S, and salt), but they also help retain water (in dry soils) or molecular O2 (in flooded soils) inside the root. More recently, outer apoplastic barriers have also been proposed to play a role in restricting greenhouse gas emissions from rice-growing areas (see this blog post). While rice stands as the predominant model plant, numerous other species exhibit the capability to alter their cell walls, consequently modulating the apoplast within the outer parts of their roots. These modifications occur in response to environmental signals and consist of depositions of biopolymers in the cell walls in the outer part of the root (namely sclerenchyma, exodermis, and epidermis.
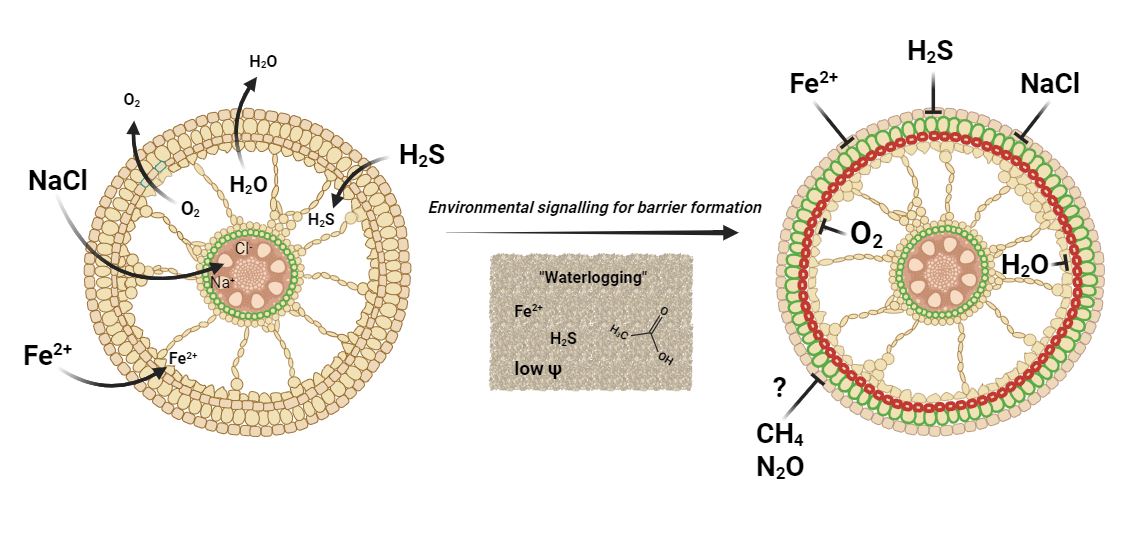
Some wetland plants form a constitutive barrier to radial O2 loss, whereas others, including rice, can form an inducible barrier in response to environmental signals. In rice, the outer apoplastic barriers are formed during soil flooding, but neither low O2, high CO2, nor elevated ethylene, which are all characteristics of flooded soils, induce the formation of outer apoplastic root barriers. Instead, low concentrations of low molecular mass organic acids (e.g., acidic, propionic, butyric, and hexanoic acids) and H2S, and high concentrations of NH4+ and NaCl all lead to swift modifications of the cell walls of the exodermis and/or the sclerenchyma. The cell wall modifications consist of suberization of the exodermis and/or lignification of the sclerenchyma, but regardless of the main biopolymer involved, apoplastic movements are affected, as indicated by low radial O2 loss, low permeance to O2 of the outer part of the root, and restricted intrusion of Fe2+, H2S, or NaCl. Recently, it has been shown that low water potential (Ψw) can also induce apoplastic barrier formation in rice, primarily through lignin depositions in the sclerenchyma.
While several environmental signals for inducible barrier formation have been identified, the hormonal and genetic regulation of exodermal suberization is poorly understood. Shiono and coworkers have previously shown that a rice mutant with a defective ABA biosynthesis gene (osaba1) exhibited no exodermal suberization and a very weak barrier to radial O2 loss, but the exodermal suberization and barrier could be recovered by supplying ABA to the root medium. Likewise, in a recent issue of Annals of Botany, Shiono and Matsuura show that exogenous application of ABA induces suberization of the exodermis in barley plants. They demonstrate that the suberization restricts radial O2 loss despite the fact that barley does not form a barrier to radial O2 loss regardless of whether it is grown in a drained or flooded soil.
Annals of Botany requested our commentary on Shiono's recent paper - our commentary is available from this link, and Shiono's paper can be accessed from here.
Some wetland plants form a constitutive barrier to radial O2 loss, whereas others, including rice, can form an inducible barrier in response to environmental signals. In rice, the outer apoplastic barriers are formed during soil flooding, but neither low O2, high CO2, nor elevated ethylene, which are all characteristics of flooded soils, induce the formation of outer apoplastic root barriers. Instead, low concentrations of low molecular mass organic acids (e.g., acidic, propionic, butyric, and hexanoic acids) and H2S, and high concentrations of NH4+ and NaCl all lead to swift modifications of the cell walls of the exodermis and/or the sclerenchyma. The cell wall modifications consist of suberization of the exodermis and/or lignification of the sclerenchyma, but regardless of the main biopolymer involved, apoplastic movements are affected, as indicated by low radial O2 loss, low permeance to O2 of the outer part of the root, and restricted intrusion of Fe2+, H2S, or NaCl. Recently, it has been shown that low water potential (Ψw) can also induce apoplastic barrier formation in rice, primarily through lignin depositions in the sclerenchyma.
While several environmental signals for inducible barrier formation have been identified, the hormonal and genetic regulation of exodermal suberization is poorly understood. Shiono and coworkers have previously shown that a rice mutant with a defective ABA biosynthesis gene (osaba1) exhibited no exodermal suberization and a very weak barrier to radial O2 loss, but the exodermal suberization and barrier could be recovered by supplying ABA to the root medium. Likewise, in a recent issue of Annals of Botany, Shiono and Matsuura show that exogenous application of ABA induces suberization of the exodermis in barley plants. They demonstrate that the suberization restricts radial O2 loss despite the fact that barley does not form a barrier to radial O2 loss regardless of whether it is grown in a drained or flooded soil.
Annals of Botany requested our commentary on Shiono's recent paper - our commentary is available from this link, and Shiono's paper can be accessed from here.